生信文章解析(第八篇)胃癌中m6A修饰的减少预示恶性表型以及Wnt/PI3K-Akt信号通路增强
m6A修饰广泛存在于哺乳动物细胞中,是mRNA以及lncRNA中最常见的表观修饰。m6A影响mRNA结构,成熟以及稳定性,从而调节蛋白质表达。然而,目前对m6A在肿瘤发生中的作用,特别是在胃癌(GC)中的作用仍不是十分清楚。今天小编就和大家分享一篇关于m6A的文章【Reduced m6A modification predicts malignant phenotypes and augmented Wnt/PI3K-Akt signaling in gastric cancer】。

在这项研究中,作者评估了m6A与GC的临床病理学相关性,并探讨了其潜在的机制。工作结果表明,低m6A信号可以预测胃癌的不良临床病理特征,并且也能够激活致癌的Wnt/PI3K-Akt信号通路,促进了胃癌细胞的恶性表型的产生。

文章收集78例弥漫性胃癌患者的组织标本(通过质谱(MS)分析和外显子组测序证实),以及来自TCGA的CG数据。

1.通过质谱分析定量m6A调控因子的表达谱
虽然有研究开发了一种基于抗体捕获的测序方法来鉴定m6A转录本,但针对患者来源的癌症标本的m6A组学研究仍然很少。为了研究m6A在GC中的作用,作者从大样本GC队列中提取了m6A调控因子的表达来表示m6A修饰的程度。并将调控因子分为三类,分别为METTL3/METTL14, YTHDF1/YTHDF2/YTHDF3, 和 ALKBH5/FTO。
2.m6A调控因子表达突变景观以及特征
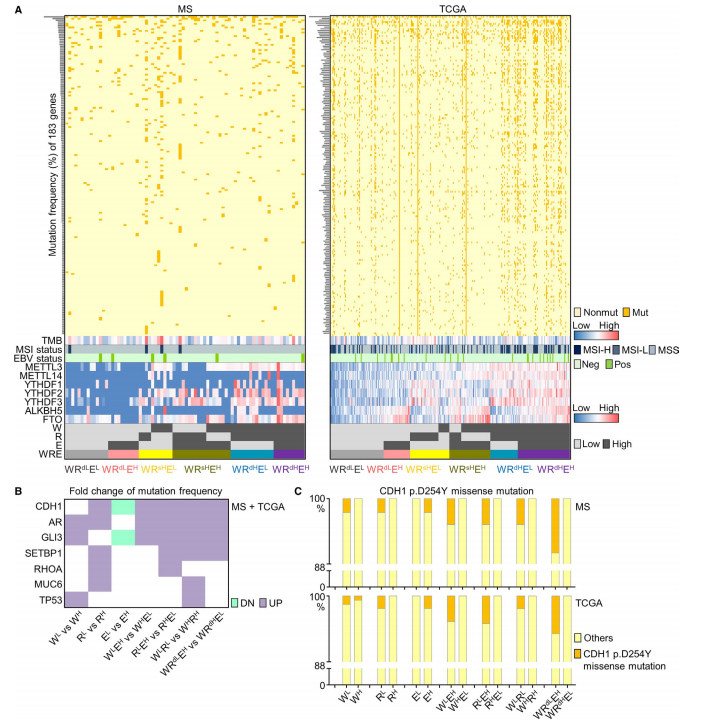 图1
图1
在这一部分,作者研究了7个m6A调控因子的分布,以及在MS队列(78例)和TCGA GC数据集中生成了3个signature(W、R、E),并评估其基因组特征的相关性。在图1A中根据表达中位数,将样本分为高表达低表达组。根据靶向外显子组测序,在MS和TCGA队列中评估关键突变,发现METTL3, METTL14、YTHDF1、YTHDF2、YTHDF3、ALKBH5、FTO均为罕见基因,MS和TCGA组中共出现了183个基因突变频率高于1%。作者接下来又在所有样本中评估了驱动基因的突变频率(图1B)。在图1C中可以看出CDH1的错义突变主要见于低m6A患者,这表明GC中的关键突变可能抑制m6A修饰。
3.m6A减少预示着胃癌的不良预后
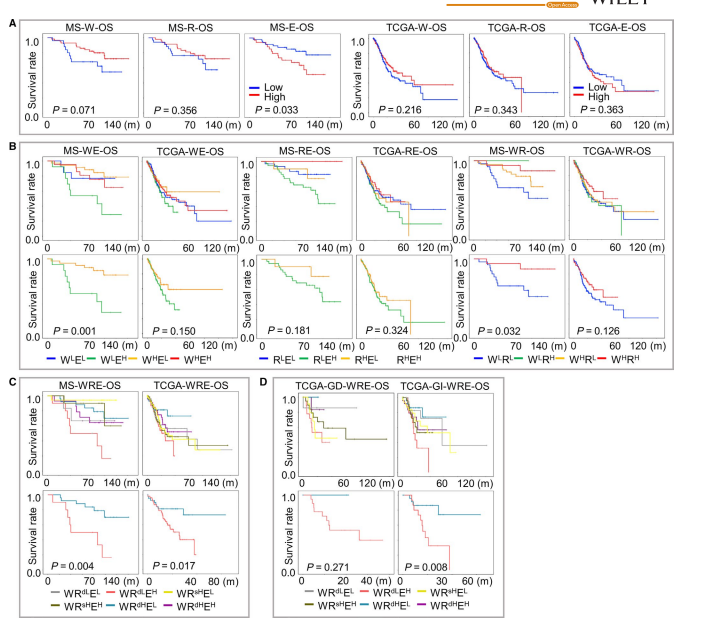
图2
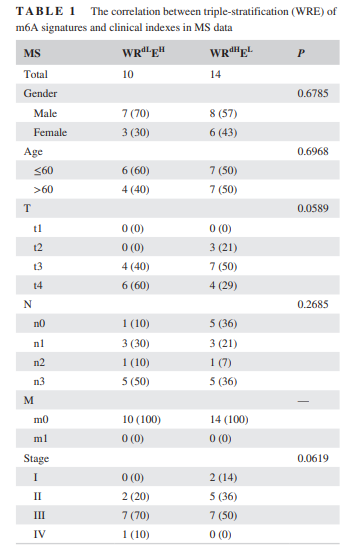 表1
表1
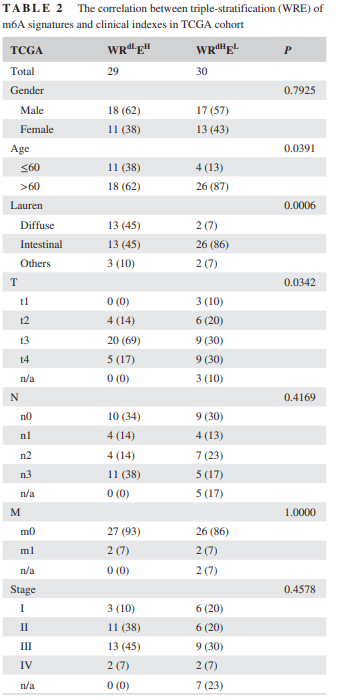 表2
表2
在这一部分,作者首先根据表达水平的中位数,将样本分为高地表达组,然后进行生存分析。图2A展示了生存分析结果。由于m6A同时受到W,R,E的调节,所以作者为了更好的反应m6A的调控,进行了两两组合的生存分析,其结果如图2B所示。而在图2C,D中可以看出,进行三重分层的生存分析结果。表1和表2分别展示了在MS以及TCGA中,m6A的WREsignature与临床指标之间的关系。
4.m6A的减少促进了GC细胞的增殖和侵袭
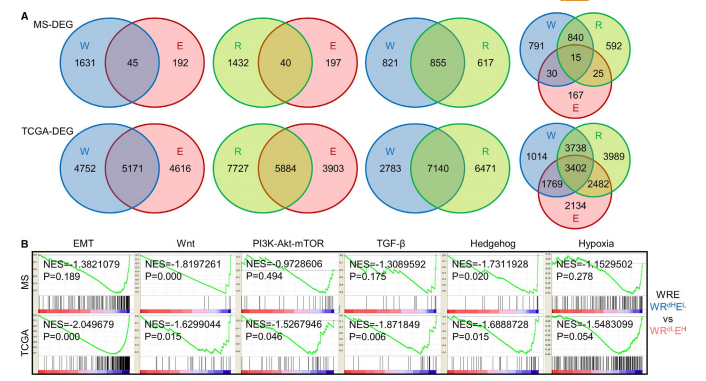 图3
图3
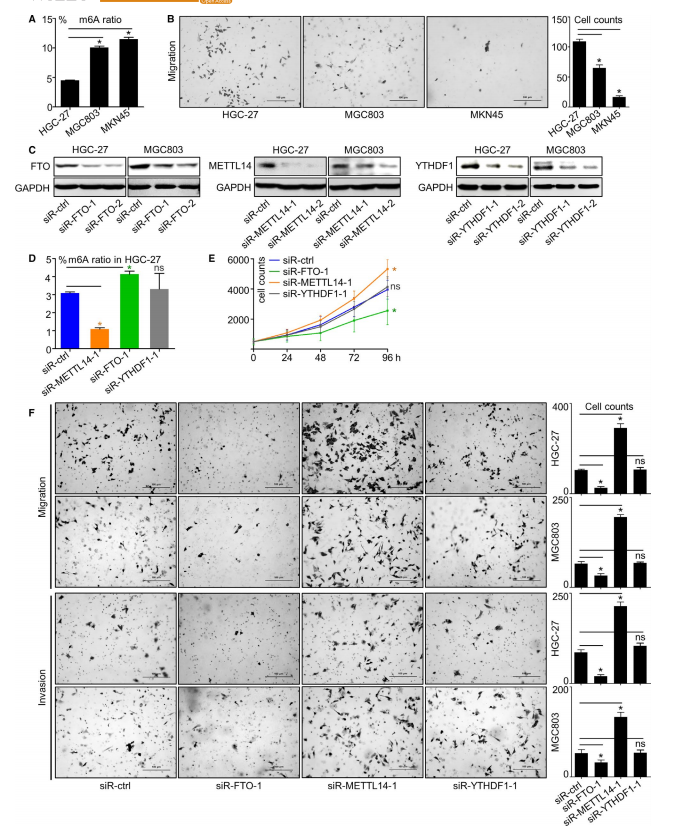 图4
图4
在这一部分作者首先进行了通路分析,来评估m6A与CG的生物学关系。图3A的韦恩图展示了高m6A特征与低m6A特征的差异表达基因情况。然后进行功能富集,功能富集结果如图3B所示。为了验证,作者评估了多个GC细胞系中m6A修饰的总水平发现在HGC-27,MGC803到MKN45带有m6A修饰的RNA的含量依次增加(图4A),迁移能力依次降低(图4B)。作者选择了3个有代表性的基因,发现这三个基因的表达被两个siRNA抑制(图4C)。图4D,E展示了进行基因敲除后的RNA以及增值率。图4F可以看出与增殖相似,METTL14敲除或FTO敲除均可增强GC细胞的迁移和侵袭能力,但对YTHDF1敲除基本不受影响。
5.m6A拮抗Wnt和PI3K-Akt信号传导
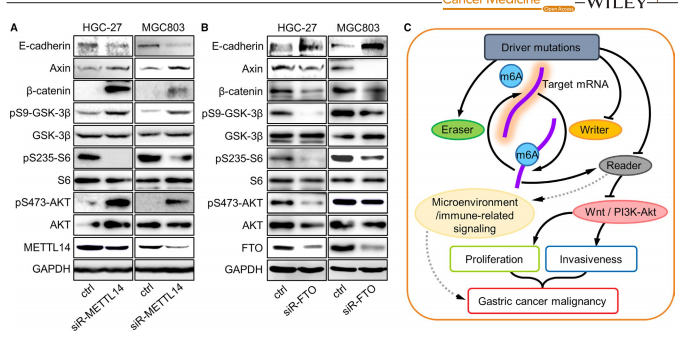 图5
图5
图5A可以看出,与GSEA结果一致,WTL和PI3K-Akt信号通过METTL14敲除激活,而图5B可以看出FTO敲除会对其抑制。接着作者构建了概述了m6A调控胃癌进展的相关网络,如图5C所示,可以看出由于干扰素在介导癌症免疫中发挥双重功能,因此m6A可通过抑制干扰素产生来调节GC的免疫应答。
到这里这篇文章的主要内容就介绍完了,总结一下,文章结合CG的表达信息,采用功能富集生存分析等方法推断出m6A-损失介导致癌信号传导(例如Wnt和PI3K-Akt)可以优先促进GC进展,系统的阐明了m6A在GC中的临床病理学作用,研究了m6A对肿瘤功能抑制及其潜在分子机制。

生信文章解析(第一篇)在乳腺癌中,MAPK通路的突变驱动扰动与肿瘤内免疫反应负相关?
生信文章解析(第二篇)使用深度学习的方法整合多组学数据预测肝癌预后
生信文章解析(第三篇)生物信息学分析膀胱癌不同亚型中免疫调节基因
生信文章解析(第四篇)能不能找到与PTC分化相关的代谢基因,且进一步识别和PTC预后相关的代谢基因?
- 发表于 2019-08-15 11:29
- 阅读 ( 9023 )
- 分类:文献解读
